|
Города как арены микроэволюционных процессов (чем обеспечивается устойчивость популяций в нестабильной, мозаичной и изменчивой среде) В. С. Фридман, Д. Н. Кавтарадзе, Г. Н. СимкинБиологический факультет МГУ, Москва В последние 50-100 лет всё новые виды птиц и млекопитающих осваивают городскую среду и формируют специализированные городские популяции. Эти процессы обычно протекают независимо в разных частях видового ареала. Чаще всего они начинаются в наиболее урбанизированных регионах и затем распространяются туда, где ландшафт и расселение людей преобразованы развивающимися городами. Достаточно вспомнить давно начавшееся формирование городских популяций больших синиц Parus major и лазоревок P. caeruleus, недавнее освоение центральноевропейских городов вяхирем Columba palumbus и рябинником Turdus pilaris, синантропизацию большеклювой вороны Corvus macrorhynchos (завершившуюся в Японии и только начинающуюся в Приморье), наконец уже вошедшую в учебники синантропизацию чёрного дрозда Turdus merula, распространявшуюся от Рейна к Днепру. Освоение одним и тем же видом разных городов, отличающихся историей застройки и соотношением основных местообитаний, протекает разными темпами и достигает разной степени завершённости. Например, резко различается степень освоения городов Европы сойкой Garrulus glandarius и сорокой Pica pica, причём течение этого процесса мало или вовсе не зависит от плотности тамошнего населения синантропных врановых (чёрной вороны, грача и галки).Во всех случаях освоение птицами городских территорий завершается появлением специализированных городских популяций. Помимо существенной специализации их экологии и поведения, давно ставшей объектом интереса натуралистов, почти все подобные популяции в демографическом отношении изолированы от населения пригородов и природных местообитаний (например, московские кряквы Anas platyrhynchos и серой вороны C. cornix, Корбут и др., 1994; Корбут, 1996). Даже если они и пополняются за счёт притока иммигрантов из естественных местообитаний (как популяции большой синицы, лазоревки или поползня Sitta europaea), это пополнение высоко избирательно - в городские популяции включаются и размножаются, видимо, только особи, соответствующие им своими эко-этологическими предпочтениями. Остальные же просто избегают высокоурбанизированных и “островных” ландшафтов, изменённых и фрагментированных человеком.Если “природные” и “пригородные” популяции обычно стабильны, пока неизменны их местообитания, большинство “городских” популяций проявляет явную склонность к экспансии и к заселению новых урбанизированных территорий (кольчатая горлица, сирийский дятел, горихвостка-чернушка, канареечный вьюрок). Важно отметить, что расселяются именно городские птицы, расселение происходит “из города в город”, минуя природные биотопы. Лишь после формирования городской популяции вида в новом месте она становится объектом иммиграции и очагом синантропизации местных птиц (чёрный дрозд на Украине и в Польше, сирийский дятел в Галиции). При подобном расширении ареала городской популяции гнездование в природных биотопах происходит лишь после становления его устойчивых поселений в городской черте: например, возникновение поселений сирийского дятла в пойменных тополёвниках и грабинниках произошло на Украине лишь через 30-40 лет после заселения городских парков. Следовательно, формирование специализированных представляет собой микроэволюционный процесс - они приобретают не только специфические отношения с городской средой (экологические и поведенческие), но и собственную историческую (в перспективе - эволюционную) судьбу , автономную от судьбы “негородских” популяций. Мы постараемся обрисовать вероятные факторы подобной микроэволюции и выявить её потенциальные механизмы, определяющие её своеобразие.Своеобразие процесса (синантропизация) и результата (специализированные городские популяции) освоения городской среды разными видами птиц и млекопитающих можно выразить “тремя слишком ”:
Поэтому мы поставили под сомнение традиционный взгляд на природу микроэволюционных процессов, сопровождающих освоение городов высшими позвоночными (см. Вахрушев, Раутиан, 1993; Мешкова, Федорович, 1996). Эти процессы обычно уподобляют формированию специализированных подвидов или популяций в новых для них местообитаниях (характерному для широко распространённых и эврибионтных видов, действительно легче прочих проникающих в города). Соответственно, формирование специфической орнито- и териофауны городов (из синантропных видов и видов-синурбанистов) представляется соприродным повторному заселению биотой ландшафта, где естественные биоценозы уничтожены катастрофой (в нашем случае техногенной). Соответственно в эволюционном аспекте (в отличие от экологического) восстановление “зональных” орнито- и териокомплексов города ничем не отличается от формирования фауны любого естественного сообщества и происходит путём специализации разных видов к осваиваемым ими новым нишам в городской среде (наиболее развёрнутую аргументацию см. Вахрушев, Раутиан, 1993). Подобная точка зрения ныне вызывает многочисленные возражения. Действительно, с трудом поддаётся обозрению и систематизации разнообразие тех обстоятельств, которые были существенны для успешного освоения каждого отдельного города каждым отдельным видом, создающих неповторимую историю каждой из городских популяций (больших синиц в Алма-Ате, чёрных дроздов в Киеве и пр.). Но все формирующиеся популяции синантропных видов и видов-синурбанистов (Мы отделяем синурбанистов от синантропных видов, поскольку они не переходят (пока?) к обитанию в районах сплошной застройки, на бульварах, скверах и иных искусственных элементах городского ландшафта. Эти виды населяют природные территории города - городские леса, луга, болота и водоёмы с естественными берегами, подвергающиеся более косвенному воздействию большинства факторов городской среды, нежели собственно городские кварталы (см. Dinetti, 1994). Например, в Москве существующие популяции кряквы напрямую связаны с местами поступления в водоёмы излишков продуктов и рассеивания излишков тепла, тогда как группировки хохлатой чернети и красноголового нырка - лишь опосредовано, через повышение продуктивности водоёмов (Авилова, 1998).) отличают одни и те же черты в их пространственно-этологической и социодемографической структуре, а также в поведении отдельных особей (прежде всего в отношении к территории и разным пятнам “мозаики” её биотопов). Эти диагностические черты “городских” популяций и особей оказываются не только универсальными, но и инвариантными:
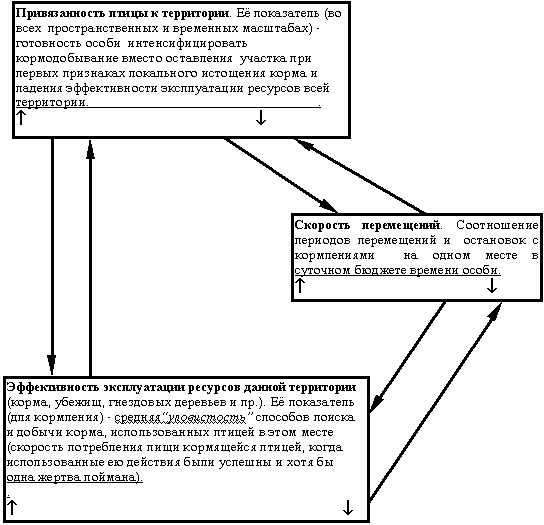
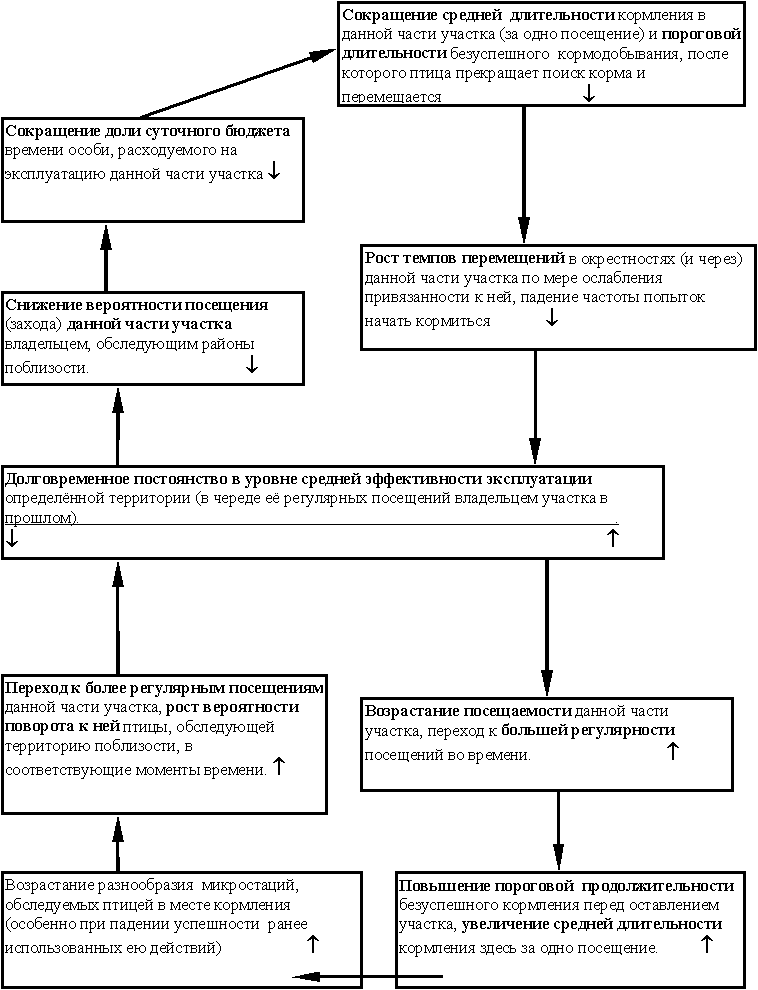
Выделенный нами инвариант популяционных и поведенческих диагностических черт вовсе не уникален для городских популяций соответствующих видов - в них он лишь наиболее облигатен. Анализ данных о существовании самых разных видов птиц в островном (для них) ландшафте (островки леса среди нелесных угодий и др.) заставляет считать, что он возникает всякий раз, когда региональная популяция сталкивается с долговременной фрагментацией своих ранее непрерывных местообитаний и вынуждена устойчиво существовать, когда её биотопы превращаются в “архипелаг” дробящихся и уменьшающихся “островков”. Соответствующий инвариант диагностических черт у видов, устойчиво существующих в “островном ландшафте”, также независим от масштаба рассмотрения “архипелага” - это и виды ядра орнитокомплексов лесных микрофрагментов в лесостепной зоне (Захарова, 2000), и виды-неурбанисты, сохраняющие долговременную стабильность своих популяций в высокоурбанизированных регионах типа Подмосковья (Фридман, Ерёмкин, 2000), и собственно устойчивые популяции на архипелагах мелких островков (Grant, Grant, 1997). Мы постараемся показать единство приспособлений птиц к обитанию собственно в городской среде и к обитанию в “архипелаге биотопов” (где фрагментация нарушила значимую для птиц непрерывность биотопа и постоянство парцеллярной мозаики его растительности), различающихся лишь по глубине и преходящести (Напрашивается мысль, что второе представляет собой начальный, ещё обратимый этап первого. Однако для её проверки в нашем распоряжении недостаточно данных.) этих изменений.Попробуем выделить и описать в явном виде те механизмы формирования синдрома диагностических черт, который диагностирует отличия специализированных городских популяций от популяций пригородов, обменивающихся с ними особями, и от близких видов, не предпринимающих попыток освоения городской среды. Наш анализ основан на данных о биологии этих популяций как в собственно городских, так и во фрагментированных и нарушенных естественных ландшафтах. Его результаты позволили охарактеризовать специфику городской среды как арены микроэволюционных процессов формирования специализированных городских популяций, неожиданно порождающих универсальный синдром взаимосвязанных приспособлений общего характера вместо ожидаемого всплеска разнообразия частных адаптаций к специфике осваиваемого видом “кусочка” городских местообитаний и частным обстоятельствам истории их заселения. Действительно, по аналогии с возникновением специализированных форм у широко распространённых видов в новых биотопах, осваиваемых широко распространёнными видами можно предположить, что по мере складывания городских орнитокомплексов степень специализации составляющих их близких видов будет возрастать, а степень перекрывания их топических и трофических ниш в городских биотопах - снижаться, как это обычно происходит в естественных сообществах (Вахрушев, Раутиан, 1993). Происходит же прямо противоположное - и у синантропных видов птиц в центральных кварталах города, и у видов-синурбанистов, населяющих более близкие к естественным биотопы (сизарь, ворона и домовый воробей, Вахрушев, Раутиан, 1993; большая синица и лазоревка в городах Европы, Dhondt, 1989 ; Toth, Tö rö k, 1994).Это противоречие заставило нас по-новому взглянуть на природу проблем, с которыми сталкиваются разные виды птиц, переходящие к регулярному гнездованию в городских биотопах. Мы перешли от подыскивания факторов адаптации, отдельных для каждого вида, к поиску универсальных механизмов (популяционных и поведенческих), позволяющих весьма различным видам птиц и млекопитающих устойчиво существовать в городской среде. Другой задачей было определение общей специфики микроэволюционных процессов, способных не только направленно формировать у них универсальный синдром диагностических черт (или же развивать изначально существовавшие потенции, Корбут, наст. сборн.), но и обеспечивать их инвариантность относительно масштабов хронотопа существования разных популяций. Природа этих эволюционных механизмов служит предметом дискуссий сторонников селективного формирования специфических черт “городских” птиц со сторонниками изначальной преадаптированности успешно осваивающих городскую среду видов (см. Мешкова, Федорович, 1996; Корбут, 1996). Обе стороны рассматривают эти экоэтологические черты в качестве адаптаций общего значения, приспосабливающих популяционную организацию и поведение особей к “островному ландшафту” городских биотопов в целом - к определённому уровню нестабильности и изменчивости очертаний значимых для птицы “островов” и к соотношению числа “островов” разной площади в “архипелаге” (см. Baglioni et al., 1998; Фридман, Ерёмкин, 2000), но не к биотопам отдельных “островов”.Однако сторонники преадаптации части видов к освоению городской среды предлагают неселекционистское объяснение быстрого формирования адаптаций общего значения в специализированных городских популяциях (Корбут, 1996). Они считают их результатом процессов самоорганизации популяционной и социодемографической структуры вида, стимулируемых развивающей средой обитания в городе, но не микроэволюционным процессом образования соответствующих адаптаций. Последний должен был бы включать дифференциальную элиминацию, крайне интенсивную по причине высокой скорости становления городских популяций, однако ни у одного из изученных видов при освоении городских территорий элиминация не была ни высока, ни (что главное) избирательна.Данные же о повышенной частоте в городских популяциях генетических, морфологических и окрасочных аберраций могут быть либо объяснены особенностями питания птиц, (Характерно избыточное количество массового корма для взрослых птиц при резком дефиците биологически наиболее ценных кормов, богатых животными белками и микроэлементами, особенно для маленьких птенцов и вылетающих слётков. Поэтому максимум частоты аберраций в окраске у домового воробья, серой вороны и других синантропных видов отмечен у слётков и в первые недели после вылета. Как показывают результаты их индивидуального мечения, в дальнейшем проявление этих аберраций (альбинизм, хромизм) у большинства особей постепенно исчезает, а сами они живут много лет и зачастую размножаются лучше птиц “дикого типа”.) либо вызваны ненаправленной дестабилизацией онтогенеза в городской среде. Наиболее частый у городских птиц тип аберраций - существенное повышение уровня флюктуирующей асимметрии в городских популяциях (и популяциях в “островном” ландшафте), которое тем больше, чем меньше по площади занимаемые “островки” в “архипелаге” биотопов ( Turdus merula, Moller, 1995). Это именно тот тип ненаправленной изменчивости, который не может быть векторизован селективным воздействием любой интенсивности. К тому же большинство аберраций окраски сравнительно с их аналогами в негородских популяциях проявляют себя полным или частичным разрушением структурных элементов окраски "дикого типа" (частичный альбинизм у домовых воробьёв и ворон, окраска "кофейной" и "черночеканной" морф сизаря), но не созданием новых.Поскольку именно преобладание адаптаций общего значения определяет для натуралиста "облик" экологии и поведения у видов-синантропов и синурбанистов, встаёт вопрос об эволюционном смысле их ускоренного формирования. Их появление не повышает приспособленности отдельных особей к существованию в городе, не обеспечивает стойкого возрастания успеха размножения в городских популяциях и почти не увеличивает устойчивости к отдельным угнетающим средовым воздействиям. В популяциях, обитающих в ненарушенных биотопах, основной жизненной стратегией является минимизация риска (Поэтому размещение гнездовых территорий у разных особей в одном и том же местообитании в значительной степени повторяется из сезона в сезон, а их выбор определяется не столько обилием, сколько доступностью корма и безопасностью кормёжки (Черенков и др., 1995).) разных форм деятельности особи (вероятности гибели кладки и птенцов, риска хищничества, беспокойства при кормлении и пр.), нежели максимизация эффективности этой деятельности. Соответственно в спектре заселяемых ими биотопов степень их предпочтения (и первоочередного заселения) возрастает обратно пропорционально рискованности долговременного существования там птиц и относительно независима от его эффективности (см. Barkan, 1990; Esa et al., 1994).В городской среде те же самые излюбленные местообитания вида часто оказываются “экологической ловушкой”, привлекательной при кратковременном посещении, но опасной для длительного существования вследствие высокого риска гибели кладки или птенцов (от бродячих кошек в кулисах кустов и пр.), концентрации тяжёлых металлов, пестицидов и иных загрязнений в техногенных рефугиумах фауны типа полей орошения и пр. (Птицы техногенных водоёмов..., 1997). Поэтому в городских популяциях успешность каждой из непосредственно наблюдаемых форм поведения - кормового, территориального, защитного и особенно брачного - существенно ниже, чем в пригородных и негородских популяциях этих же видов ( Dhondt, 1989; Ксенц, 1990; Esa et al., 1994) из-за повышенной частоты экзогенных прерываний соответствующей деятельности. Особенно заметно снижение эффективности брачных демонстраций и постоянства пар в популяциях синантропных видов, при одновременном превращении интенсивного и сезонного ухаживания в вяло текущее почти в течение круглого года (Ксенц, 1990; Корбут, 1994).Возникающие адаптации общего значения оставляют почти неизменными отдельные элементы, слагающие видоспецифические стереотипы экологии и поведения особи: репертуар демонстраций и стереотипных способов поиска/добычи корма, организация цепочек стереотипных действий у демонстрирующей птицы при ухаживании или угрозе, у кормящейся - при обследовании участка, специфические тактики поведения в разных ситуациях. Однако становление этих адаптаций вызывает направленные изменения в закономерностях сочетания и комбинаторики таких элементов (характеризующих системные свойства экологии популяций и поведения индивидуумов у соответствующего вида). Подобные правила переключения и комбинаторики стереотипных видов деятельности птицы в быстро и часто меняющейся обстановке позволяют своевременно и адекватно регулировать деятельность особей в подобной ситуации. В “негородских” популяциях это чаще всего происходит при необходимости адекватного разделения времени на разные формы деятельности птицы в ответ на быстрые изменения контекста социального общения (предъявление соседом территориальных претензий самцу, в этот момент ухаживающему за самкой, см. Фридман, 2000), в “городских” и “островных” - из-за необходимости постоянно оптимизировать выбор разных форм поведения в ответ на постоянное появление новых опасностей и новых возможностей в разных частях их участков (Ксенц, 1990; Корбут, 199 4, 1996; Baglioni et al., 1998; Мешкова, 2000а, б). Исследования синантропных популяций серой вороны и кряквы, скворца и чёрного дрозда, сизарей, больших синиц, “островных” популяций чёрных дроздов и мухоловок-пеструшек показали неизменное сохранение тонкой экоэтологической специфики вида и в городских популяциях, которые любой натуралист сочтёт сверхспециализированными. Однако в них сохраняется не только состав, но и тип распределения частот тех жизненных стратегий и тактик существования, которые характерны для вида(Примером альтернативных жизненных стратегий служат “помоечники” и “летуны” у сизаря. Первые всю свою жизнь проводят в том дворе, где расположена колония, а перемещения происходят лишь во время дисперсии молодняка (обычно не далее соседней колонии). Вторые регулярно летают кормиться на пригородные поля или элеваторы, равно как легко переселяются даже в достаточно далёкие колонии. (Ксенц, 1990). Полиморфизм поведенческих тактик в городских популяций отражается в частотах предпочтений гнездовых деревьев с разной архитектурой кроны, разной породы, разной частоте использования антропогенных материалов в конструкции гнезда у гнездящихся по соседству пар и пр.) - сменяются лишь доминирующие стратегии/тактики и параметры частотного распределения (у сизаря и серой вороны, см. Ксенц, 1990; Корбут, 1996). Именно универсальные приспособления в “городских” и “островных” популяциях создают возможность гораздо большей привязанности размножающихся пар и резидентных особей к их многолетним участкам обитания (у нетерриториальных видов - зонам активности) (Вообще постоянство территориальных связей у резидентных особей/размножающихся пар в городских и “островных” популяциях много выше, чем вне города (и в “материковых” местообитаниях, сравни резкое возрастание территориальной и биотопической привязанности в финской и подмосковной популяциях D.leucotos по сравнению с обитателями Беловежской пущи, Virkkala et al., 1993; Wesolowski, 1995; Фридман, 1998). Наоборот, она резко уменьшена у эпизодически и вовсе неразмножающихся особей из “популяционного резерва” - помощников, нетерриторриальных сателлитов, холостующих не только в “городских”, но и в “островных” популяциях вполне урбофобных видов. У того же D.leucotos резко возросшая подвижность диспергирующего молодняка и холостующих птиц при возрастании территориального консерватизма пар эффективно препятствуют воспроизводству сокращающейся популяции вида, населяющей “островки” северных мелколиственных лесов среди хвойной монокультуры (Virkkala et al., 1993).) В границах таких участков оказывается заключено наибольшее разнообразие "кусочков мозаики" из разных биотопов, эксплуатируемых владельцем (Sukopp, Trepl, наст. сборн.). Эти биотопы существенно различаются между собой “для резидента” по уровню эффективности и риска при эксплуатации локальных запасов корма, укрытий, гнездопригодных деревьев и пр. ресурсов; в естественных же биотопах при сопоставимой мозаичности растительности степень функциональной дифференциации таких “фрагментов” (по значимости для долговременного существования резидента) территории на порядок ниже. В естественных биотопах долговременное обитание и успешное размножение пар определяются в основном интегральным качеством их территории - её кормовой ценностью и ненарушенностью (большой площадью массивов при меньшей инсуляризации) предпочитаемых видом типов растительности (например, Eckert, Weatherhead, 1987a, b; Moller, 1995). Наоборот, в городских и “островных” ландшафтах для птиц наиболее значимо дифференциальное качество территории - наличие отдельных ключевых элементов, служащих сигналом о возможности долговременного обитания (или регулярного кормления, как это делают расселяющиеся D. minor на московских бульварах и во дворах), даже если давно разрушено большинство элементов первоначальной пространственной структуры парцеллярной мозаики растительности. Их наличие позволяет птицам устанавливать прочные территориальные связи и успешно размножаться даже внутри городских дворов и в центре мегаполиса. Примеры: старые деревья на опушке рядом с плоскими крышами гаражей для московских горихвосток, старые тополя для мухоловки-пеструшки и лазоревки, голубые ели для зеленушки (см. Симкин, 1988; Михайлов, 1991). В силу высокой гетерогенности и изменчивости мозаики биотопов на участке птицы, условием их долговременного обитания там (как и обитания всей местной популяции в ареале внутри “островного” ландшафта) становится долговременное же постоянство средней эффективности и риска эксплуатации резидентом одних и тех же ресурсов в данной части участка (в череде её последовательных посещений). Такое постоянство позволяет постоянно обитающим здесь птицам существенно снизить уровень риска при исполнении наиболее стереотипных форм социального и повседневного поведения, повысив “предсказуемость для них” среды и контекста их осуществления, одновременно облегчая адекватный выбор последующих форм поведения птицы и разделения времени между ними (тем более что эффективность, риск и стереотипность исполнения прямо пропорциональны друг другу у всех изученных форм видоспецифического поведения - угрожающих и ухаживательных демонстраций, оборонительных действий и действий поиска/добычи корма). Исходя из вышесказанного, наилучшим показателем устойчивости существования особи или популяции на определённой территории будут вероятностные, а не детерминистские критерии, отражающие возможности деятельности скорее, нежели её текущий успех (вероятность реализации репродуктивного потенциала, а не успех размножения). Можно предложить 2 серии подобных критериев устойчивости для “городских” и “островных” популяций: 1) индикаторы риска существования особи/популяции в её хронотопе (вероятность гибели прежних обитателей участка, их смены в ходе территориальной конкуренции, оставления участка из-за затяжного ухудшения условий существования и пр.); 2) индикаторы способности уменьшать риск до “природного” уровня за счёт своевременной реактивности (у особей - поведенческой, у популяций - за счёт изменений структуры расселения и направлений дисперсии). Такими критериями могут быть успех размножения за все сезоны абонирования участка, вероятность сохранить привязанность к участку и его биотопам и при резком (затяжном) ухудшении условий существования за счёт освоения на тех же территориях новых ресурсов корма, убежищ, мест гнездования, районов поиска партнёра, вероятность успешного сопротивления территориальным претендентам и пр. Частота таких случаев существенно повышена в “городских” и “островных” популяциях (Михайлов, 1991; Мешкова, Федорович, 1996; Мешкова, 2000). Однако детальное исследование “негородских” популяций позволяет обнаружить потенциальную способность к такому “нетипичному” поведению, правда с меньшей частотой реализации (Корбут, 1996). Часто столь “нетипичные” способы кормления, гнездования, коллективной защиты в городских условиях оказываются вовсе не недавним приобретением, но филогенетически древней и наследственной чертой предковых популяций вида или всей группы видов; их реализация - результат сигнальной индукции соответствующими местообитаниями (Симкин, 1988, 1995). Примером служит групповое гнездование чёрных коршунов в азиатских городах, многих видов хищных птиц в лесополосах и лесных микрофрагментах в лесостепи (Захарова, 2000), колониальное гнездование малого зуйка Charadrius dubius на топком дне спущенных рыборазводных прудов (Симкин, наст. сборн.). При становлении “городских” и “островных” популяций у самых разных видов максимизируются показатели устойчивости второго рода, отражающие всё большую способность долговременных резидентов участка использовать постоянство уровня эффективности и риска своей деятельности в одних и тех же его частях для экстраполяции ожидаемого успеха (и риска) деятельности там в будущих посещениях (также Мешкова, Федорович, 1996; Бадридзе, 1998). Даже особи из “диких” популяций разных видов в эксперименте демонстрируют способность к подобной экстраполяции “в будущее” долговременно-устойчивого уровня эффективности кормления, благодаря чему повышается суммарное число жертв, добытых ими в определённой части участка (но не уловистость единичного кормления, Nishimura, 1988). Успешное приспособление к городской среде требует от птиц и млекопитающих именно способности экстраполировать подобные долговременные тенденции “в будущую деятельность” (привязывая их к определённому хронотопу и “отсеивая” флюктуации от направленных изменений), при недостаточности простой реакции на изменения среды, даже самой быстрой и точной. Слишком уж высока степень изменчивости и нестабильности отдельных “мозаик” местообитаний на участке животного (даже в аналогах природных биотопах - лесных, болотных и пр. “островках”); обстановка там меняется ото дня ко дню настолько существенно и быстро, что развёртывание видоспецифического поведения особи (повседневного и социального) требует сохранения преемственности последовательных стереотипных действий в долговременно развёртывающихся видах деятельности (угрожающих и брачных демонстраций при образовании пары, стереотипов кормодобывания при обследовании участка), нежели быстроты/адекватности изменений в выборе очередного действия (в ответ на постоянные изменения в обстановке и в окружении действующей птицы). И кормление, и ухаживание, и охрана территории - контекстуально-зависимые и контекстуально-управляемые виды деятельности (хотя её элементы и представляют собой инстинктивные реакции) (см. Панов, 1978). Поэтому в изменчивой и нестабильной среде большая быстрота и точность “ответных” изменений, вносимых животным в выбор следующих действий при высокой стереотипности их исполнения начинает “работать против птицы”. Увеличивая эффективность каждого отдельного действия (добычи корма, угрозы захватчику, демонстрации ухаживания) в конкретной части участка, она снижает способность особи сопрягать разнородные действия в направленно развёртывающуюся долговременную деятельность - ухаживание за периодически прилетающим партнёром, охрану участка от проникающих захватчиков, устойчивую эксплуатацию изменчиво распределённых кормов. Развёртывание такой векторизованной деятельности почти автономно от экзогенной стимуляции даже в случае территориальной или брачной коммуникации, а её долговременный успех определяется преемственностью в сменяющих друг друга реакциях (по уровню их мотивации и изменениям эффективности и риска), хотя каждая реакция функционально специфична для процесса общения или же кормления.Действительно устойчивость существования особей в городских и “островных” ландшафтах обычно прямо пропорциональна полноте и точности реализации видовых стереотипов их поведения в мозаике биотопов участка вопреки её высокой изменчивости, нестабильности и гетерогенности среды (то же относится к местной популяции и её хронотопу). Реактивные же, приобретённые и пластичные формы поведения, как это ни странно, играют подчинённую роль при обитании в мозаичной и нестабильной среде. Во всяком случае они неэффективны вне устойчивых связей особи с определённым хронотопом и мозаикой его местообитаний. Подобные связи обычно стабилизируются при росте социального ранга резидентов (по отношению к соседям по группировке) и “срока их пребывания” в составе группировки, при одновременном сокращении риска дестабилизации этих связей. И то, и другое происходи за счёт избирательного исчезновения из локальных поселений именно тех особей, чей социальный статус, уровень метаболизма и активности в разных видах взаимодействий “отклоняется” от названной пропорциональности ( P.major, Гаврилов, Керимов и др., 1996; Dendrocopos major, Friedmann, 1998).Можно сделать вывод, что для обитателей городских (и “островных”) ландшафтов сохранение филогенетической косности имеет преимущества перед проявлением селективной гибкости. В результате несколько снижается адекватность адаптивного ответа особей на кратковременные изменения условий существования, но резко повышается устойчивость сопряжения надындивидуальных параметров структуры их популяций (включая развёртывание видоспецифических и стереотипных форм поведения) с “медленными” процессами долговременного развития городской среды. Именно к их неблагоприятному течение не могут адаптировать виды, избегающие проникать на городские территории - для них это “медленные катастрофы”, не угрожающие текущему существованию, но снижающие устойчивость воспроизводства популяции. Такая ситуация является результатом действия на структуру популяций и паттерны поведения индивидуумов стабилизирующей, а не движущей формы отбора. Она увеличивает защищённость долговременного существования особи/популяции от “шума” краткосрочных флюктуаций городской среды в постоянном хронотопе. Обратная картина характерна для популяций в ненарушенных биотопах - их жизнеспособность определяется именно оппортунистичностью поведения и социодемографической структуры локальных поселений, быстро изменяющихся в ответ на изменения качества территорий, благоприятности сезона размножения и пр. (рис.1 ).Если малонарушенные лесные сообщества и городские территории обладают в общем сходным (высоким) уровнем мозаичности, то огни кардинально отличаются характером динамики этой мозаичности во времени. В первом случае соотношение разных “островков в архипелаге” (разных парцелл растительности внутри характерной площади выявления) остаётся стабильным, если только темп появления “окон” древесного полога (разного размера, вплоть до зубровых полян) соответствует темпу восстановления в них видов древесных эдификаторов: эта динамика благоприятна для постоянно гнездящихся здесь видов птиц (Смирнова, 1998; Преображенская, 1999). Наоборот, на городских территориях (и шире - в урбанизированных регионах в целом) соотношение “островков разных местообитаний в архипелаге” направленно меняется. (Эти тенденции характерны и при рассмотрении больших масштабов урбанизированных территорий, например, всего "архипелага" естественных местообитаний в регионе типа Подмосковья. Разрастание центральной агломерации постоянно увеличивает мозаичность этого "архипелага " за счёт фрагментации "островов" естественных экосистем (и "отплывания" их друг от друга в "архипелаге"). Её темпы снижаются на градиенте от урбанизированного центра к сохранившейся периферии региона, при возрастании средней площади и падении дистанции между сохранившимися "островами" природных сообществ. Поэтому градиент изменений "островного ландшафта” в пространстве от центра к периферии региона одновременно указывает на устойчивый вектор преобразований мозаики его экосистем во времени (Фридман, Ерёмкин, 2000).) Эти тенденции неблагоприятны для населяющих их видов: чем крупнее и менее нарушен “остров”, тем выше средняя скорость сокращения его площади и вероятность его фрагментации на более мелкие “островки” (Фридман, Ерёмкин, 2000). Поэтому особи из городских (и “островных”) популяций, успешно существующие в подобной среде, вынуждены максимизировать консерватизм своих территориальных и биотопических “насколько возможно”, пока устойчиво их существование в данном хронотопе, - при сохранении возможности почти мгновенного перехода к предельной их лабильности после потери участка (рис.1 ).В городском и “островном” ландшафте выживание отдельных особей и жизнеспособность всей популяции не требует специальных адаптаций к конкретным изменениям среды - и сформированных ещё до её освоения, и возникших de novo. Однако безусловно необходимо ускоренное (относительно прочих приспособлений) развитие адаптаций общего значения. Последние позволяют абонентам долговременных участков обитания приспосабливать свою экологию и поведение (а устойчивым популяциям - характер расселения вида, тип динамики численности и миграционные потоки между поселениями) к определённому уровню мозаичности, изменчивости и нестабильности городской среды per se тем точнее, чем долгосрочнее их связь с соответствующим хронотопом.(Здесь опять уместны понятия теории стабилизирующего отбора. Его основным результатом оказывается рационализация онтогенеза и рост его помехозащищённости, которая снижает вариабельность морфологии конченой стадии последнего существенно сильнее, чем элиминация уклоняющихся вариантов (Шишкин, 1988). При формировании же специализированных городских популяций и у синантропных видов, и у видов-синурбанистов возрастает помехозащищённость процессов развёртывания видоспецифических (наиболее стереотипных) форм поведения (у особей) и поддержания пространственно-этологической структуры в популяциях.) Следствием преобладания таких адаптаций становится повышение системного уровня организации поведения "городских" птиц и их популяционной структуры за счёт создания возможностей опережающего отражения и заблаговременного прогноза развития ситуации, позволяющих не разделять, а сопрягать во времени и пространстве поддержание устойчивости "стержневых" поведенческих стереотипов и своевременное "перенацеливание" процесса их развёртывания адекватно изменениям в обстановке и окружении действующей птицы. Даже успех процессов образования пары и изгнания претендентов на территорию в городских популяциях определяется в процессе долговременной персонализации отношений между соседствующими резидентами и более зависит от способности социальных партнёров к долговременной синхронизации повседневного поведения в пространстве участков, нежели от успешного воздействия брачных и территориальных демонстраций и ритуализованных форм конкуренции в этих сферах. Подобные изменения весьма характерны для всех популяций, существующих в островном ландшафте ("island effect”, Hurst, Gray, 1998), но наиболее выражены в городских. Здесь даже возможность успешно образовать пару и размножиться определяется не столько эффективностью взаимного ухаживания будущих партнёров per se, сколько устойчивостью существования на их собственных территориях, влияющей на регулярность их встреч и ухаживаний в течение длительного периода (гораздо более растянутого). Поэтому приспособленность их членов определяется большей или меньшей возможностью реализации репродуктивного потенциала, и мало зависит от текущего успеха их размножения и факторов, обычно влияющих на этот успех в природных популяциях (сроки начала яйцекладки, интенсивность и частота копуляций перед яйцекладкой, качество гнездового участка и пр.), см. Ксенц, 1990; Esa et al., 1994. В совокупности, стратегия существования птиц в природных популяциях оппортунистична: она реактивно "подстраивается" к изменениям в эффективности кормления, успехе размножения и другим показателям эффективности существования особи/популяции на определённой территории. Жизненная стратегия городских (и “островных”) популяций ригидна: выбор тактики эксплуатации ресурсов в разных частях района обитания регулируется экстраполяцией (на будущие посещения) прошлого уровня эффективности этой эксплуатации в меру его долговременной устойчивости (рис.1) Сказанное означает сдвиг у "городских птиц" соотношения К /r-стратегий в сторону преобладания К (а в природных популяциях - r), хотя экология и поведение вида не меняются. Этот феномен остаётся необъяснённым в рамках традиционных воззрений на особенности микроэволюционных процессов, формирующих специализированные городские популяции.В самом деле, пресс специализированных хищников в городах почти исчезает, а количество и доступность корма много выше. Возрастание смертности у взрослых особей (а особенно гибели гнёзд и выводков) на городских территориях по традиционным взглядам на соотношение r- и К-отбора должно сдвигать популяцию в сторону r-стратегии воспроизводства, с высокой скоростью потребления ресурсов (с низкой эффективностью их потребления и усвоения), трансформирующихся в основном в репродуктивное усилие, а не в поддержание устойчивости популяционной организации (Розенберг и др., 1999). В результате растут и удельная скорость размножения, и смертность, но первая - ускоренно, а вторая - линейно.Однако в городских популяциях птиц и млекопитающих отмечена прямо противоположная картина - рост средней продолжительности жизни (и длительности абонирования постоянных участков), рост консерватизма территориальных и биотопических связей не только у размножающихся территориальных пар, но и у скапливающихся на их участках нетерриториальных "сателлитов" и “помощников”, при снижении среднего успеха размножения за сезон в таких "постоянных" популяциях (см. Ксенц, 1990; Корбут, 1996 ; Baglioni et al., 1998). Этот же феномен вызывает существенное уменьшение остроты конкуренции за территории у “городских” птиц, равно как и готовности совместно обитающих особей разного статуса вступать друг с другом в любые ритуализованные антагонистические взаимодействия, в том числе и для оспаривания статуса конкурента и повышения собственного (см. Hurst, Gray, 1998).Разнообразие страт максимально именно у долгоживущих “городских” птиц: размножающие и холостующие владельцы постоянных участков, постоянно обитающие здесь нетерриториальные птицы, регулярно появляющиеся мигранты, пытающиеся завладеть частью участка, члены кочующих стай находятся в постоянном контакте. Для этих особей характерны и другие черты, сопряжённые с вышеназванными: рост взаимной терпимости птиц, в том числе и у тех, которые в “природных” популяциях обречены своим статусом почти исключительно на конкурентные и антагонистические отношения - соседние территориальные пары или соседние холостые самцы, поющие на постоянных участках, при свободном перемещении между их участками самок, ещё не образовавших пары. Одновременно отмечается существенное учащение дружественных и игровых взаимодействий между животными одного статуса даже в ситуациях наиболее острой территориальной или сексуальной конкуренции, причём и у птиц, и у млекопитающих. Новизна же тех ситуаций, в которые постоянно попадают "городские" животные, гораздо слабее инициирует проявления неофобии и оборонительных реакций, но интенсифицирует исследовательскую активность (см. Мешкова, Федорович, 1996; Котенкова, Мешкова, 2000). Частота и интенсивность территориальных стычек с соседями и конфликтов с мигрантами намного выше именно в городских популяциях (в том числе и сравнительно с близкими видами-урбофобами)(В “городских” популяциях резко вырастает доля нерезидентных особей, в каждый данный момент времени ищущих новый участок для долговременного обитания, а также доля неразмножающихся птиц из “популяционного резерва. Выше и скорость/дальность перемещений таких особей. У большинства видов в ареале городских популяций регулярно зимуют особи из пригородов и из городов, расположенных севернее-восточнее (большая синица, поползень, серая ворона, грач, большой пёстрый дятел).), но ожесточённость и ритуализованность таких стычек гораздо ниже, а победа и поражение в них “гораздо менее бесспорны”.Невозможность присутствия побеждённой птицы на участке победителя в городских и “островных” популяциях гораздо менее вероятна даже при постоянных поражениях, нежели в более разреженных поселениях в ненарушенных биотопах (Симкин, 1977 ; Hurst, Gray, 1998). Казалось бы, большая скученность животных на гораздо меньших "островках" подходящих местообитаний будет вызывать обратные эффекты. Однако они отмечены лишь у видов, вполне сосуществующих с человеком, но отступающих из городов на следующем этапе их урбанизации и разрастания (грач, галка и пр.). Результат таких стычек также парадоксален - это быстрый рост терпимости к постоянному присутствию соседей и птиц других страт вместо ожидаемого обострения нетерпимости. Если бы формирование городских популяций было обычным селективным процессом (который эффективен именно при обострении конкуренции и использовании всех возможностей для немедленной максимизации репродуктивного усилия), то обнаруженный “эффект островной терпимости” неизбежно расширял бы возможность размножения у нетерриториальных или подчинённых птиц, что сдвигало бы схему воспроизводства популяции к r-стратегии.Однако ничего подобного не отмечено. Хотя численность популяции и плотность её населения по мере освоении города возрастают экспоненциально, одновременно все динамические и лабильные характеристики в её экологии и поведении сменяются консервативными (как у “допущенных” к размножению владельцев долговременных участков, так и в среднем по популяции). Любопытно, что совершенно аналогичное противоречие между “ r-типом динамики численности” и “К-типом” социодемографической структуры популяции отличает и человеческую популяцию.(Хорошо известно, что предложенная Мальтусом модель экспоненциального роста N(t )=ert неприложима к популяциям большинства животных и растений из-за существования значимой отрицательной корреляции между N и r. Однако в популяциях человека эта корреляция отсутствует по крайней мере в последние 300 лет после начала массовой урбанизации (даже до промышленной революции).)Но как же тогда обеспечивается быстрый рост городских популяций большой синицы, чёрного дрозда, серой вороны, кряквы, вяхиря, рябинника и других подобных видов, осваивающих современный развивающийся мегаполис (существенно отличающийся даже от промышленного города в начале века, пространственная структура которого - центр, промышленные окраины, слободы, была весьма консервативна и менялась плавно, Корбут, наст. сборник)? Ответ на этот вопрос даёт идущее одновременно резкое возрастание доли особей, в данный момент нетерриториальных или негнездящихся (к которым владельцы участков, достаточно жёстко защищаемых от соседей, относятся на порядок терпимее, чем к птицам из популяций пригородов). Эти особи постоянно обследуют зону своей активности, у них повышено внимание к любым изменениям обстановки, способным облегчить приобретение долговременной территории при повышенной же толерантности к людям, машинам и прочим “безразличным к ним” явлениям городской жизни.(Сходные изменения в поведении зарегистрированы и в случаях, когда в городской черте много лет существуют регулярные зимовки данного вида (хохлатые чернети и красноголовые нырки в ФРГ, казарки на побережье Голландии и пр.).) Поскольку активность и лабильность (в том числе и в социальных контактах) этих особей низкого статуса повышена, а не подавлена, как в популяциях , населяющих природные биотопы (см. Eden, 1985; Birkhead et al., 1986), благодаря крайней подвижности их биотопических и территориальных связей (Особенно важно, что перемещения у подобных особей совершаются в том же самом хронотопе, в котором обитает и воспроизводится местная популяция, и лишь “при отсутствии вакансий” они его оставляют. В природных популяциях картина противоположна: возрастание подвижности и активности подобных особей всегда ведёт к их переселению в другой район (см. Eckert, Weatherhead, 1987a, b).) локальные популяции на городских и “островных” территориях способны своевременно и эффективно использовать все те возможности для закладки новых участков обитания и успешного размножения на них, которые постоянно создаются локальными изменениями среды в пределах ареала.В популяциях вне города у большинства видов птиц переселение на новый участок или в новое поселение происходит лишь после неуспешного сезона размножения (его дальность обычно обратно пропорциональна сокращению успеха размножения особи сравнительно со средним по поселению). В городской среде сами по себе невозможность размножения или риск гибели на участке обитания или поблизости ничуть не индуцируют переселений у резидентов и не снижают стремления птиц из "популяционного резерва" завладеть соответствующей территорией и стать резидентами самим. Падение же успеха размножения и даже полная гибель гнёзд в одном сезоне обычно не снижает прочности дальнейших территориальных и биотопических связей владельца со своим участком. Однако постоянно происходящие локальные изменения мозаики городской среды и её биотопов вызывают своевременные и адекватные перераспределения особей из "популяционного резерва", быстро заселяющих и успешно гнездящихся во всех вновь возникающих "островках" подходящих местообитаний. В городских популяциях репродуктивный потенциал каждой отдельной пары в каждом из сезонов заметно снижается, но гораздо выше возможность его полной реализации за жизнь особи. В сочетании с ростом средней продолжительности их жизни это позволяет видам-синантропам и синурбанистам быстро увеличивать свою численность в городах. Возможно, сходный механизм ответственен за нынешний стремительный рост населения (именно городского) в развивающихся странах, а в прошлом веке - и в промышленно развитых. Ведь максимальная репродукция - 16-18 выживших детей у одной женщины за жизнь - достигается лишь в популяциях австралийских аборигенов, с трудом просто восстанавливающихся после регулярных засух. Резкий же рост населения начался сразу после превращения городов из центра торговли и ремёсел в центры промышленной и особенно политической активности (ранее бывшие в усадьбах землевладельцев) и проходил в условиях падения среднего числа выращенных детей на семью и устойчивого падения фертильности у обоих полов (числа жизнеспособных сперматозоидов в эйякуляте, рост частоты невынашиваний беременности, резкое снижение вероятности зачатия и пр.). Следовательно, изменения городской среды вообще не могут выступать в качестве факторов адаптации, к которым приспособлены экология и поведение городских популяций высших позвоночных, и которые ответственны за появление у них специализации к городскому образу жизни (она, видимо, просто отсутствует). Они выступают лишь в качестве факторов риска (как любые неспецифические нарушения, время от времени неожиданно увеличивающие гетерогенность и изменчивость “мозаики” ландшафта, но не меняющие значения отдельных её “фрагментов). Приспособление к ним требует не новых специальных адаптаций, а большей “помехозащищённости и “точности наведения” от механизмов популяционной и индивидуальной устойчивости, способных и при высокой рискованности действий птицы (или слагающегося из них риска для существования местной популяции) обеспечить направленное развёртывание видоспецифических стереотипов её кормового, социального, исследовательского и пр. поведения. В этом случае деятельность особи может протекать при высокой предсказуемости изменений в мозаике её хронотопа (при обитании в ненарушенных естественных биотопах её создаёт высокой постоянство паттерна их ценотической структуры). Лишь в этом случае сиюминутные действия животного будут с высокой вероятностью адекватны долговременным целям её деятельности (аналогичная адекватность у человека обычно называется смыслом, сравни, Братусь, наст. сборн.).Рис.1. Схема регуляции жёсткости территориальных и биотопических связей в природных и городских (“островных”) биотопах (показаны известные положительные обратные связи между явлениями)А. Члены поселений в высокомозаичных естественных местообитаниях - оппортунистические изменения территориальных привязанностей вслед за изменениями в эффективности эксплуатации разных частей участка.
Обратите внимание, что в случае А территориальные и биотопические связи птиц изменяются под давлением внешних для птицы изменений в кормности "островков" разных местообитаний, а в Б эти изменения носят существенно более эндогенный и независимый от текущих изменений характер - связь с территорией превращается в привязанность к ней благодаря возможности воздействия пластичного поведения птицы на управляющий параметр - долговременное постоянство средней эффективности эксплуатации той или иной части участка (в А это, очевидно, невозможно). Поэтому в Б ослабление связи птицы с одними частями территории увеличивает долговременное постоянство эксплуатации других частей участка, привязанность к которым соответственно возрастает Б. Высокомозаичные городские и “островные” местообитания (мозаичность которых возникает в процессе инсуляризации и фрагментации массива, а не возобновления парцеллярной структуры его растительности) - ригидное охранение прежних территориальных привязанностей, пока эффективность эксплуатации разных частей участка остаётся долговременно стабильной и предсказуемой при будущих посещениях, и быстрый переход к полному отсутствию связи с территорией в нестабильной и изменчивой ситуации
Вахрушев А.А., Раутиан А.С. Исторический подход к экологии сообществ// Журн. общ. биол. 1993. Т.54. №.5. С.532-553. Гаврилов В.М., Керимов А.Б., Голубева Т.Б. и др. Популяционный и экологический эффект стай и групповых поселений на примере большой синицы [in English]// Орнитология. Вып.28. М.: изд-во МГУ, 1998. С.52-59. Захарова Н.Ю. Население птиц лесных микрофрагментов Восточно-европейской лесостепи. Автореф. и дисс. Канд. биол. наук. М.: 2000. 173 с. Корбут В.В. Питание. Особенности жизни кряквы; Изменчивость городских птиц// Урбанизированная популяция водоплавающих (Anas platyrhynchos) г.Москвы. М.: Аргус, 1994. С. 89-150. Корбут В.В. Уникальная популяция серой вороны г.Москвы. Депрессия численности серой вороны в Московском регионе// Доклады РАН. 1996. Т.348. №.1. С.136-139, 140-142. Мешкова Н.Н., Федорович Е.Ю. Ориентировочно-исследовательская деятельность, подражание и игра как психологические механизмы адаптации высших позвоночных к урбанизированной среде. М.: Аргус, 1996. 226 с. Михайлов К.Е. Опознание гнездовых ситуаций и пусковые механизмы расселения у птиц// Современная орнитология. М.: Наука, 1991. С.5-21. Котенкова Е.В. Мешкова Н.Н. Феномен синантропии: приспособление синантропных видов домовых мышей и становление синантропного образа жизни в процессе эволюции// Животные в городе. Мат. Научн.-практ. конф. М.: 2000. С.115-117. Ксенц А.С. Колониальность у сизого голубя: диалектика консерватизма и лабильности// Современные проблемы изучения колониальности у птиц. Симферополь-Мелитополь: СОНАТ, 1990. С.33-37. Мешкова Н.Н. Пищедобывательное поведение серой вороны в г.Москве. Поведение бездомных собак на территории Московского метрополитена// Животные в городе. Мат. Научн.-практ. конф. М.: 2000а. С.124-126, 127-130. Птицы техногенных водоемов Центральной России. М.: изд-во МГУ. 150 c. Симкин Г.Н. Групповое поселение большого пестрого дятла// Орнитология. Вып.13. М.:изд-во МГУ, 1977. С.134-145. Симкин Г.Н. На пути к разработке новых стратегий охраны и экологической оптимизации природной среды// Бюлл. МОИП. 1988. Т.93. Вып.1. C.11-23. Фридман В.С., Ерёмкин Г.С. Поляризация ландшафта как фактор риска для региональных авифаун центра России// Сохранение биоразнообразия и рациональное использование биологических ресурсов. Тез. докл. I молод. Школы. М.: 2000. С.105. Baglioni V., Marcos J., Bogliani J. Offprint, dispersal, cooperative breeding and habitat saturation in two crow populations// Variation in infraspecific behaviour. - Abstr. ASAB Summer Meet., Urbino, 1998. Р.13-14. Barkan Ch. A field test of risk-sensitive foraging in black-capped chickadees (Parus atricapillus)// Ecology. 1990. Vol.71. №1. Р.391-400. Birkhead T., Clarkson K. Ceremonial gathering of the magpie Pica pica: territory probing and acquisition// Behaviour. 1985. Vol.94. №3-4. Р.324-332. Dhondt A. Ecological and evolutionary effects of intgerspecific competition in tits// Wilson Bull. 1989. Vol.101. №2. Р.198-216. Eckert Ch., Weatherhead P. Ideal dominance distribution: a test using red-winged blackbirds (Agelaius phoeniceus)// Behav. Ecol. and Sociobiol. 1987a. Vol.20. P.43-52. Eckert Ch., Weatherhead P. Owners, floaters and competitive asymmetries among territorial red-winged black-birds. Anim. Behav. 1987b. Vol.35. №5. Р.1317-1323. Eden S. The social organisation of non-breeding Magpies Pica pica// Ibis. 1989. Vol.131. №1. Р.141-153. Esa H., Jukka J., Pekka R. Nest site selection in pied flycatcher Ficedula hypoleuca and forest fragmentation// Contr. 21st Int. Ornith. Comgr. J. Ornithol. 1994. Bd.135. Sonderheft. S.197. Friedmann W.S. Signal-used mechanism generated diversity of the alternative strategies in territorial behaviour and they consequences for social structure steadyness in Dendrocopos major settlements: a field study// Intraspecific variations in behaviour. - Abstr. ASAB Summer Meet., Urbino, 1998. - P.32-33. Hurst J., Gray S. Behaviour underlyiung the “island syndrome”// Variation in infraspecific behaviour. - Abstr. ASAB Summer Meet., Urbino, 1998. Р.42. Moller A. Developmental stability and ideal despotic distribution of blackbirds in a party environment// Oikos. 1995. Vol.72. №2. Р.228-234. Nishimura K. Foraging behaviour of ural owls (Strix uralensis) in a patchy environment: the impotance of acquired information// Ecol. Res. 1988. Vol.3. №3. Р.319-332. Toth L., Tö rö k J. Asymmetric competition for food between great and blue tits during the breeding season// 1994. Contr. 21st Int. Ornith. Comgr. J. Ornithol. 1994. Bd.135. Sonderheft. S.181. Virkkala R., Alanko T., Laine T., Tiainen J. Population contraction of the White-Backed Woodpecker in Finland as a consequence of habitat alternation// Biol. Conserv. 1993. Vol.66. P.47-53.
|